Abstract
Rodents in the family Heteromyidae are subject to hypercapnic and hypoxic conditions in burrows. This experiment dealt with Heteromyids and their reactions and ventilatory responses to increasing concentrations of CO2. Two genera and four species of rodents were tested, with a minimum of three different individuals of each species for the tests. Initially they were tested and observed for behavioral responses to CO2 increases of 2% at 5-minute intervals. Another experiment was conducted to measure their ventilatory response to 5% CO2 increases every 3 minutes. All of the rodents showed some sort of behavioral response to levels of CO2 below 15% (calculation in torr). They did not leave the burrows at higher concentrations even though they had access to fresh air outside the burrow. When we measured their ventilatory responses to increasing levels of CO2, we found that their minute ventilation increased until the CO2 levels reached 15% (sea level adjusted). We concluded that Heteromyids are able to compensate for increased levels of CO2 up to 15%, which is the maximum sustainable level.Introduction
The medullary respiratory center in mammals depends upon carbon dioxide (CO2) for the regulation of breathing. Since atmospheric air contains approximately 0.03% CO2, these respiratory centers are very sensitive to increased levels of CO2. Fossorial rodents, in the wild, spend much of their time in burrows. This results in a hypercapnic environment, which is characterized by high concentrations of CO2. Burrow levels of CO2 have been reported to be between 3% and 9.5%, depending upon the species and burrow depth (Tenney and Boggs, 1986). This would present a problem to non-fossorial mammals, but does not do so in adapted animals. Chronic exposures to hypoxic (low concentrations of O2) and hypercapnic environments are known to lead to adaptive changes in ventilation and ventilatory responses (Dempsey and Forster, 1982), some of which are due to changes occurring at the level of the carotid body chemoreceptors (Nielson et al., 1988).
When CO2 levels increase, blood pH decreases and chemoreceptors in the medullary center send a signal to increase ventilation. It is established that fossorial rodents do have an increase in minute ventilation in response to increasing CO2 levels, although the response curve is not as steep as in humans (Figure 1), (Tenney and Boggs, 1986). An elevated blood buffering capacity is a contributing factor in allowing a burrowing rodent to tolerate moderately high levels of CO2 (Tenney and Boggs, 1986). By increasing the plasma bicarbonate, blood-buffering capacity increases and blood pH returns to normal. Therefore the signal to increase ventilation ceases and ventilation returns to a level only slightly higher than normal despite the elevated level of CO2 (Randall et al., Animal Physiology, 1997). Burrowing rodents have lower ventilatory responses to increased CO2 levels than do surface dwellers (Prange et al. 1998). Prange et al. found that desert rodents showed no preference for simulated burrows with ambient air over burrows containing 5% CO2.
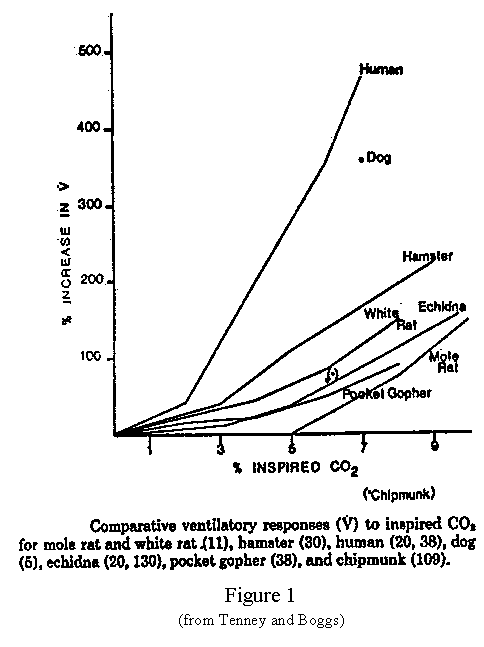
The purpose for the first part of this study was to observe and record the behavior of the fossorial rodents, Heteromyids, at increasing levels of CO2 and to determine at what point they would refuse the hypercapnic environment. The goal was to determine at what level of Pco2 they would no longer voluntarily remain in a burrow.
The goal of the second part of the study was to quantify the ventilatory response of Heteromyids to increasing CO2 levels.
Hypotheses:
- Fossorial rodents should have a maximum level of CO2 that they will voluntarily tolerate. If burrow CO2 levels are continually increased, at some point they should abandon the burrow.
- There should be a notable ventilatory increase in response to increased partial pressures of CO2 (Pco2).
- The increase in minute volume in relation to increasing CO2 levels of Heteromyids will be less than that of non-fossorial mammals, but greater than that of completely fossorial rodents such as the pocket gopher and the mole rat.
Question:
Will there be statistically significant differences in tolerance and compensation to increased levels of CO2 in different species and/or genera of fossorial rodents? If there are differences, will there be a correlation between the masses of animals and their willingness to tolerate increased levels of CO2?
Materials and Methods
Experimental AnimalsWe captured five different species of rodents using Sherman live-traps set in various locations around Owens Valley, Inyo County, California. The experiment was conducted during a reproductive period, so only males were brought back to the lab for testing. All of the rodents were housed in a laboratory at White Mountain Research Station, located in Bishop, California. They were housed in a room with windows, so that they experienced a normal light dark regime. They were housed separately in either the simulated burrows or in stainless steel cages. Both were filled with local sand and those not housed in the simulated burrows were given PVC tubing for shelter. Room temperature was around 20°C. Diets consisted of mixed seeds supplemented with fruit slices and greens.
Part I:
Eleven animals from two genera and four species were tested in this
portion of the experiment. We created the simulated burrows using glass
terrariums, PVC pipe or Habitrail tubes for tunnels, sand,
polyvinyl tubing for gas supply and measurement, duct tape, and
see-through plastic bottles as the observation/burrow chamber.
The observation chambers were held to the glass with enough duct tape so
that the sand that was applied over them could not interfere with
observation. The terrarium bottom was lined with empty plastic bottles
and cans and then a layer of sand placed over that, preventing the
terrarium from being too heavy to move. The animals were given from
several hours to a day to adjust to their new "home" before
being tested. A large amount of seed was placed inside the burrow chamber
and a small amount of sand to eliminate any foraging needs. Animals that
did not stay in the burrows for long periods of time were not used for
the behavioral study.
The voluntary tolerance test was conducted by using compressed nitrogen, oxygen, and carbon dioxide. The gas levels were administered using a GF-3 Gas Mixing Flowmeter (Cameron Instrument Company) and pumped into the chamber with polyvinyl tubing to the rear of the burrow chamber (furthest away from the tunnel entrance). Burrow CO2 (Bco2) levels were monitored with another permanently placed polyvinyl tube and extracted using 60 mL syringes and run through a CD-3A Applied Electrochemistry CO2 analyzer and CO2 sensor (Ametek corporation). The analyzer was calibrated using the Gas Mixing Flowmeter. When Bco2 levels exceeded the 16% analyzer capacity, the samples were diluted using a 1:1 dilution and the readings doubled.
The total gas mixture was pumped into the burrow starting at 100 CC/minute beginning with concentrations of 6%. The concentrations were increased by two percent at five-minute intervals until the source concentration reached 12%, at which time they were increased by two percent at ten-minute intervals. The maximum level of administered CO2 varied with each animal. The Gas Mixing Flowmeter was limited to a maximum level of 10 CC/minute so that in order to administer increasing amounts of CO2 the total flow was decreased with each increase above ten percent CO2 (Table 1). CO2 and nitrogen (N2) concentrations were varied while oxygen (O2) was always maintained above 160 torr, at a constant concentration of 32.7%. A permanently placed tube was used to monitor actual burrow concentrations of CO2 (Bco2) at regular intervals. This was done to correspond to either behaviors or to increases in administered gas concentrations.
|
(percent) |
(cc/min) |
(cc/min) |
(cc/min) |
(cc/min) |
|---|---|---|---|---|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Flow concentrations used during Part I of the experiment (not adjusted for sea level)
Any apparent behavior or visible physiological responses that were observed during the testing periods were noted as to time and Bco2 concentration. If they exited the burrow, they were gently coaxed back in with paper rustling or a gentle nudge. A decision was made prior to experimentation that if they could not be easily coaxed back into the burrow then it would be evaluated as a refusal and the experiment stopped. The animals were not closely monitored following the experiment, but were observed periodically.
Part II:
We measured ventilation using a flow-through whole-body plethysomograph
(Malan, 1973; Jacky 1978). A mixture of gas flowed simultaneously through
a reference chamber and the flow-through chamber containing the rodent.
Ports on the side of the flow-through chamber and the reference chamber
connected tubing to either side of a differential pressure transducer
(Validyne, model DP103-18). This arrangement isolated the chamber
transducer from ambient pressure noise. Incurrent and excurrent air
through both chambers passed through flow resistors. These flow resistors
were constructed by fitting pieces of PE 10 tubing (Clay Adams, no. 7401)
in 3mm i.d. polyvinyl tubing (Tygon, #3603), (Cameron, 1986). The lengths
of PE tubing (range 1to 3 cm) in each resistor were adjusted to balance
the total resistance in each circuit and between the chambers. They also
smoothed the flow through the chambers, providing a low noise pressure
signal. The pressure transducer signal was demodulated (Validyne, model
CD-15) and sent to a chart recorder (Gould, model 2400S) for
amplification and low-pass filtering. The processed ventilation signal
was also sent to the computer, which used a software routine to detect
ventilation peaks and then stored them concurrent with the metabolic rate
data. (as modified by Szewczak et al, in preparation). The gas mixture
entering the chamber was analyzed for percent of CO2 and O2
using CD-3A Applied Electrochemistry CO2and O2
analyzer and sensor (Ametek Corporation).
A total of six rodents, three from each genus were tested. The rodents were placed in the flow-through chamber, and lowered into a water bath in order to keep a constant temperature of 25°. A baseline ventilation was established on each animal without any CO2 being administered. CO2 levels were then increased at 4% intervals, up to a maximum of 24%. The level of O2 in the gas mixture was kept above 25% prevent a hypoxic environment. Following the testing, a size D battery or a rock the approximate size of the animal was placed in the chamber for calibration recordings. A syringe on top of the flow-through chamber with a volume of 200uL was expelled into the chamber and the corresponding pressure difference was recorded as above. 10-15 calibrations were run and then the average value of those readings was used to calculate ventilatory volumes of the rodents.
Results
Part I:We found that there was no significant difference between species and genera and their apparent tolerance to hypercapnic conditions. All of the rodents showed some sort of behavioral change at a level much lower than they ultimately tolerated. Six of the eleven were noted as having left their burrows as their initial response to increased CO2 levels. The others had behavioral changes that were interpreted as a discomfort to the increasingly hypercapnic environment. The mean value at which there was a behavioral change was 11% CO2 (9.5%, sea level adjusted) (Figure 2).

The most significant behavior in this study was that at the highest levels of Bco2, all of the animals remained in the burrows. The tests were stopped for one of two reasons, either we were unable to significantly increase the Bco or the level was so high that we were concerned about the animal's health. Above 15% Bco2 we noted that there was a decrease in responsiveness by all of the animals. Attempts to get their attention by tapping on the glass were also not as effective or in some cases had no effect at higher concentrations. All of the subject animals showed a very noticeable increase in ventilation. Three of the animals showed visible decreases in ventilation at Bco2 concentrations higher than 20%. Four of the six Perognathus tested showed shivering responses at Bco2 concentrations higher than 20%. This response was not noted in the Dipodomys. Overall, the Dipodomys were calmer than the Perognathus. Maximum supplied CO2 levels ranged from 24% to 28%. The maximally attained levels of Bco2 ranged from 18.78 to 26.16% (Table 2).
Table 2
|
|
|
|
(per cent) |
(per cent) |
(per cent) |
(per cent) |
|
||||
|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
|
|
|
||||
|
|
D. merriami |
|
|
|
|
|
|
|
|
|
|
|
|
D. panamintinus |
|
|
|
|
|
|
|
|
|
|
|
|
P. longimembris |
|
|
|
|
|
|
|
|
|
|
|
|
P. longimembris |
|
|
|
|
|
|
|
|
|
|
|
|
D. merriami |
|
|
|
|
|
|
|
|
|
|
|
|
D. panamintinus |
|
|
|
|
|
|
|
|
|
|
|
|
P. longimembris |
|
|
|
|
|
|
|
|
|
|
|
|
D. merriami |
|
|
|
|
|
|
|
|
|
|
|
|
P. parvus |
|
|
|
|
|
|
|
|
|
|
|
|
P. parvus |
|
|
|
|
|
|
|
|
|
|
|
|
P. parvus |
|
|
|
|
|
|
|
|
|
|
LB: left burrow
1/2T: went halfway up tunnel (Counted as leaving burrow)
BA: Behavioral Change
HTE: head in tunnel entrance
SH: shivering response
We were unable to determine a definitive, quantitative response at the highest levels of CO2. It was at that point that we decided to proceed to the second part of the experiment to quantitatively determine the respiratory response due to increased levels of CO2
Part II:
The respiratory frequency of the 6 rodents tested increased until about
15% CO2 at which time the frequency started to drop (Figure 3).
The smallest rodent, Perognathus longimembris, had the greatest
increase in respiratory frequency whereas the largest rodent had the
smallest increase in respiratory frequency. Even after the respiratory
frequency started dropping at 15% CO2, the frequency still
remained above the baseline value.

The tidal volume of each breath remained steady or only showed a slight increase among the genus Perognathus. The rodents from the genus Dipodomys showed and increase in tidal volume until about 15% CO2 at which time it started dropping rapidly (Figure 4). The Dipodomys merriami actually had a smaller tidal volume than at the beginning of the experiment, demonstrating a relative deterioration since at 15% CO2 the frequency and the tidal volume of the breaths decreased.
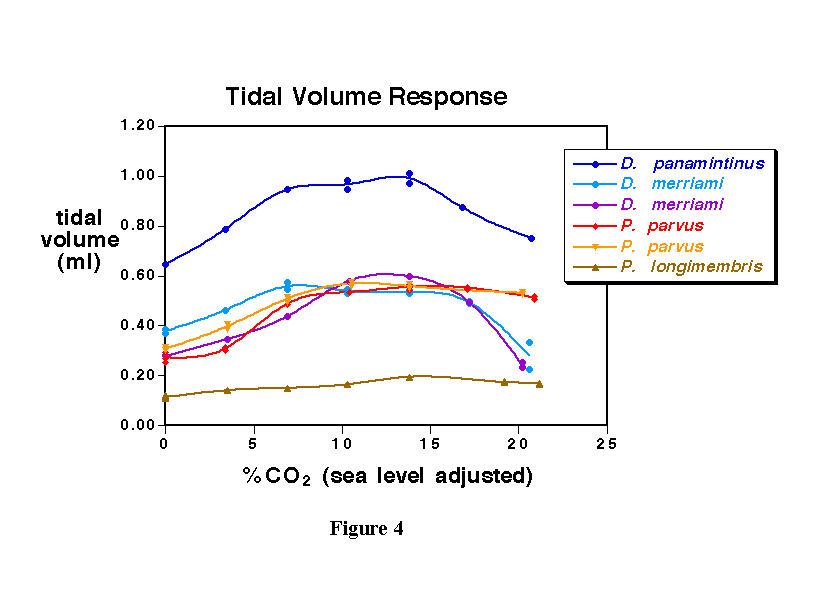
Similar to what was seen in the previous two tests, the increase in minute volume for all of the rodents dropped off at 15% CO2 (Figure 5). By 20% CO2, the species of Dipodomyshad fallen to near normal levels of minute ventilation, whereas the Perognathus appeared to be able to sustain the minute volume at a much higher level.

Discussion
Part I:The experiment began with an attempt to behaviorally quantify a maximum level at which Heteromyids could no longer be coaxed into tolerating higher and higher levels of CO2. Most animals were easily coaxed back into their burrow after exiting, while others never left the burrow. For those that did exit the burrow and were coaxed back in, they eventually quit exiting. We observed rapid breathing at CO2 levels above 11%, but eventually saw breathing drop off to lower rates. During testing it appeared that the smaller animals were able to tolerate higher levels of CO2 but when we analyzed the data and recorded behaviors, this was not the case. We concluded that when the animals exited the burrow, even as an initial reaction, that they were reflecting intolerance to the CO2 levels. Further increases in the CO2 levels seemed to incapacitate them. The animals all appeared to recover differently. Most had increased respiration for several hours, while others showed rapid recovery. One notable example was the final P. parvus, which following exposure to 26% CO2, was digging and eating seeds within three minutes of the CO2 removal. In further experiments the recovery time should also be documented, for at least thirty minutes. It would be particularly useful if respiration were monitored quantitatively during that time.
Controls:
In this portion of the experiment we attempted to use a non-fossorial rodent, Mus musculus, as a control. However, even after allowing several days for him to adjust to the simulated burrow, he never went into the burrow. Hence, for Part I, no control existed other than allowing for an adjustment period to the burrows. We did not use two D. merriami due to their failure to utilize the burrows sufficiently for the purposes of the experiment. One of these two was used in Part II and he had the highest increase in minute ventilation. So the only animals that were used were ones that utilized the burrow. Under normal conditions, without additional CO2, the animals used remained in the burrows. The only times that they were noticed outside the burrows was during the quiet hours of the night. This was detected by observing their tracks and/or signs of digging in the sand. However, a better control for this would have not only been to find non-fossorial animals that would utilize a simulated burrow (or design an environment that could still test their voluntary tolerance to increased CO2 levels), but to test the same animals with ambient air pumped into the burrows and characterize their responses.Equipment:
The largest problem with this experiment was a lack of uniformity in the simulated burrows. The burrows were made for different sized animals, without consideration for decreased gas flows. The tunnels could be longer with at least one turn in them. This would minimize CO2 loss due to air currents and assist in building and maintaining a higher concentration more rapidly. It would also be better to have enough time and burrows to allow the animals a longer period of adjustment to their surroundings. Some of the animals had enough sand in their burrows to use it to block the tunnel entrance allowing for a larger concentration of CO2 to build up. This happened towards the end of the testing and would have been very helpful had all of the animals had the same option.
We were left perplexed by our observations and decided to evaluate their actual respiratory responses to elevated CO2. We had some questions, most of which could not be answered in the scope of this course. We wondered if there was some physiological coping mechanism taking over at some point allowing them to tolerate the high CO2 levels, or if their bodies were just too overwhelmed by the conditions to allow them to try to leave the detrimental environment. It would have been helpful to be able to sample blood gasses as well as blood buffer concentrations. The literature told us that fossorial rodents do have a greater ability to acclimatize to higher levels of CO2 than do non-fossorial rodents. However, we rejected this as a reason for the animals remaining in the burrows. We observed animals that appeared to be in distress. We knew that there were respiratory changes occurring, but we did not feel confident to characterize them solely through observation. We were certain that we did not see animals recovering or less stressed at any time during the experiment. Therefore the conclusion that they had not increased levels of blood buffers to deal sufficiently with the increased CO2, was an easy one to make. The disturbing problem in attempting to interpret the behavioral study was that we could not quantify the animals� respiratory response to the increasing CO2 levels. At that point we decided to take the study further and quantify respiratory responses.
Part II:
This experiment allowed us to put together the behavioral observations
with quantitative data and helped us to better hypothesize as to what was
occurring as result of the higher levels of CO2. We believe
that the animals were able to compensate for the increasing levels of
CO2 by an increase in respiratory responses. Tidal volume and
the number of breaths per minute increased, resulting in an increase in
minute volume. The ventilatory response increased up to approximately 15%
CO2 at which point a decrease in ventilatory response began
and continued to decrease, but remained above normal levels (figure 4).
We concluded that at 15% CO2 (sea level adjusted) was the
maximum sustainable level for the tested animals. We hypothesize that at
that point their blood buffering capacity is no longer able to efficiently
deal with the excess CO2. We needed both the behavioral
observations and the quantitative data to arrive at this conclusion. The
animals had approximately four times as long to acclimatize to each
increase of CO2 in the behavioral study. Hence, the most
likely reason that the rodents did not leave the burrow at the higher
levels of CO2 was due to incapacitation.
We feel that as fossorial rodents, they do have a blood buffering capacity, but that CO2 tolerance is still limited. This study was conducted over short periods and did not allow for long term acclimatization. This study has some shortfalls in that only eleven rodents were tested in part I, with three or fewer of each species, in part II only six rodents were tested, three from each genus. If more animals could be tested and the experimental protocol refined, the results would be much more significant. Furthermore, the study should be carried out with more genera and even from other orders of other fossorial rodents such as Order Insectivora and Order Lagomorpha.
Conclusion
The rodents tested in this study clearly had an incredible tolerance to high levels of CO2. Furthermore, the rodents did abandon the burrow when faced with increasing levels of CO2. By leaving the burrows they actually did show us what their maximum tolerable level was. Although they tolerated higher levels of CO2 than what they showed us by their initial responses, we believe that they would not have been able to survive those levels, even without further increases. The second portion of the study gave us the results we needed to conclude that beyond 15% CO2 (sea level adjusted), the animals would not have survived. We are not able to determine how long they could sustain that level and have no desire to do so experimentally in the laboratory.Future Research
This project was envisioned as something entirely different than what it turned out to be. It was meant to be a study of ground squirrels in an area called Horseshoe Lake, near Mammoth, California. The recent volcanic activity in the Mammoth Mountain area is believed to have resulted in the seepage of increased CO2 into the ground. Studies were conducted to determine this after thirty acres of trees died off (http://www.395.com/dead) Reports have been made that suggest that there are still fossorial rodents living in the area. Whether there are still survivors or not, is unknown as the snow pack has prevented further study in the area. Our intention was to trap rodents from that area and measure the CO2 concentrations in their burrows and compare that to other fossorial rodents.This project hopefully will only be a preliminary to studies at Horseshoe Lake and other such comparative studies with animals in their natural environment.
Acknowledgements
I am especially grateful to Joe Szewczak for his endless support. Without Joe's help and equipment this project would not have been realized. I will always remember and try to model Joe's extraordinary patience, teaching, and mentoring. I also want to thank Mark Chappell and Frank Powell for their guidance and suggestions. I greatly appreciate the help of my classmates for great moral support. I can not thank my partner in this project, Stephanie Settimi, enough. Her ideas made this project much more complete than it ever could have been without her. Thanks Steph! Most of all thanks to UCR for giving me the grant that enabled me to come and do research at White Mountain Research Station.Literature Cited
Cameron, J.N. 1986. Principles of Physiological Measurement. Academic Press, Orlando, FL. pp. 278Dempsey, J. A., and H.V. Forster 1982. Mediation of Ventilatory
Adaptations. Physiology Review Volume 62: 262-346.
Jacky, J. P., 1978. A Plethysmograph for Long-term Measurements of
Ventilation in Unrestrained Animals. Journal of Applied Physiology
45 (4):644-647.
Malan, A. 1973. Ventilation Measured by Whole Body Plethysmography in
Hibernating Animals and Poikilotherms. Respiration Physiology
17:32-44.
Randall, D., Burggren,, W., and French, K. 1997. Animal Physiology
. 4th edition. W.H. Freeman and Company. New York.
Nielsen, A. M., G. E. Bisgard and E. H. Vidruk. 1998. Carotid
Chemoreceptor Activity During Acute and Sustained Hypoxia in Goats.
Journal of Applied Physiology Volume 65: 1796-1802.
Prange, H. D., W. M. Reilly, and B. Pinshow, 1998. Voluntary Tolerance of
High Levels of CO2 by Desert Rodents. The FASEB Journal,
Abstracts PartI, Volume 12, Number 4, A419.
Tenney, S.M. and D. F. Boggs 1986. Comparative Mammalian Respiratory
Control. In Handbook of Physiology, Section 3: The Respiratory
System, Vol. II: Control of Breathing, Part 2; edited by N.S. Cherniack
and J.G. Widdicombe. Bethesda, MD: American Physiological Society,
pp. 833-855.
Webb, Cheryl L. and William K. Milsom, 1994. Ventilatory Responses to
Acute and Chronic Hypoxic Hypercapnia in the Ground Squirrel.
Respiration Physiology. Volume 98: 137-152.
Internet website: http://www.395.com/dead (Page no longer available)
